As every individual, therefore, endeavours as much as he can both to employ his capital in the support of domestic industry, and so to direct that industry that its produce may be of the greatest value; every individual necessarily labours to render the annual revenue of the society as great as he can. He generally, indeed, neither intends to promote the public interest, nor knows how much he is promoting it. By preferring the support of domestic to that of foreign industry, he intends only his own security; and by directing that industry in such a manner as its produce may be of the greatest value, he intends only his own gain, and he is in this, as in many other cases, led by an invisible hand to promote an end which was no part of his intention. Nor is it always the worse for the society that it was no part of it. By pursuing his own interest he frequently promotes that of the society more effectually than when he really intends to promote it.
Adam Smith, An Inquiry into the Nature and Causes of the Wealth of Nations (1776)
Cité par Edwards dans son article « The fundamental theorem of natural selection » (Biol. Rev. 1994), qui cite également un propos de Sober (The nature of selection, 1984):
The Scottish economists offered a non-biological model in which a selection process improves a population as an unintended consequence of individual optimization.
Edwards mentionne également ce qu’il appelle le théorème du taux de croissance:
In a subdivided population the rate of change in the overall growth-rate is proportional to the variance in growth rates.
Ce qui donne en français: dans une population subdivisée, le taux auquel le taux de croissance global change est proportionnel à la variance des taux de croissance de chaque sous-population.
Ce théorème élémentaire est applicable partout tant qu’il y a des taux de croissance différentiels, par exemple en biologie ou en économie. Et en reliant taux et variance, il donne une piste pour comprendre le théorème fondamental de la sélection naturelle dû à Fisher. Mais tenons-nous en ici à démontrer ce théorème du taux de croissance.
Considérons une population subdivisée en 

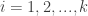

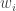

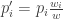
Le nouveau taux de croissance global 



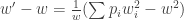
Soit finalement:

Le numérateur de cette expression correspond exactement à la variance des
Ce théorème, bien que simple, est important car il exprime clairement l’idée selon laquelle plus les taux de croissance sont différents entre sous-populations, plus rapidement les sous-populations à forte croissance vont dominer les autres. In fine, la prédominance de la plus rapide sera totale, toute la variabilité disparaîtra et le taux de croissance globale n’augmentera plus. Un peu comme si l’évolution « consommait » la variabilité.
Et ce qu’a fait Fisher, c’est « simplement » adapter ce théorème à la génétique des populations, le présentant comme l’un des résultats les plus fondamentaux de toute la biologie. Affaire à suivre…
